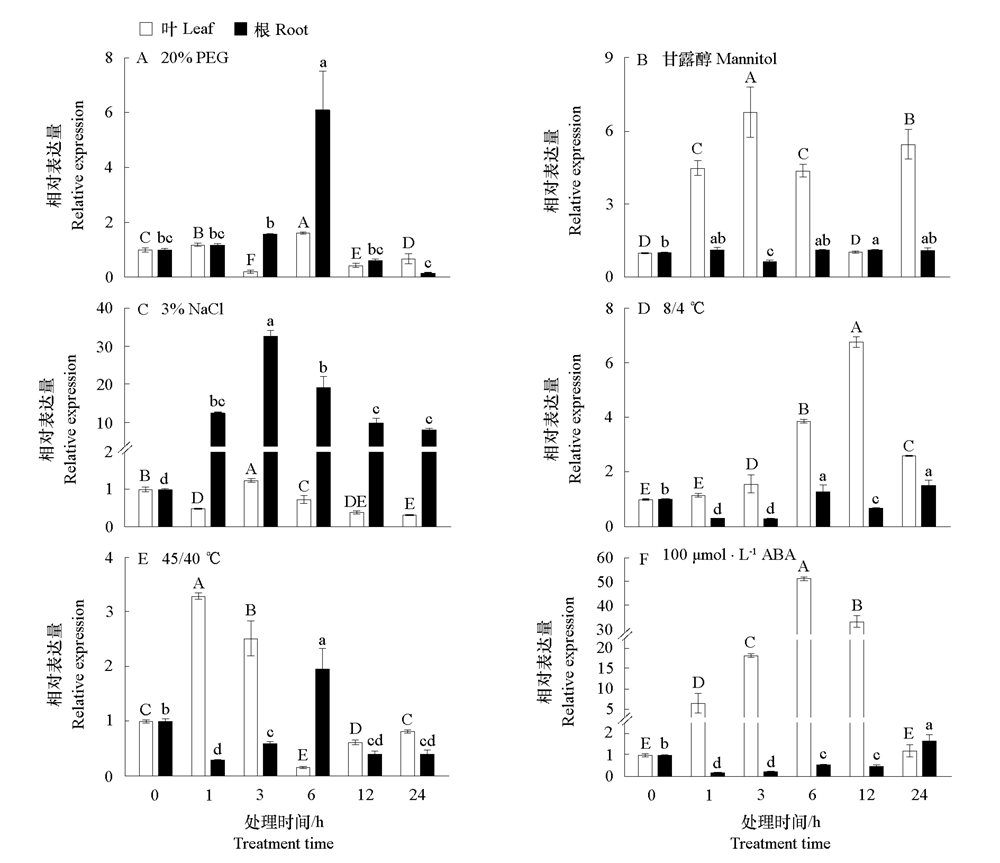
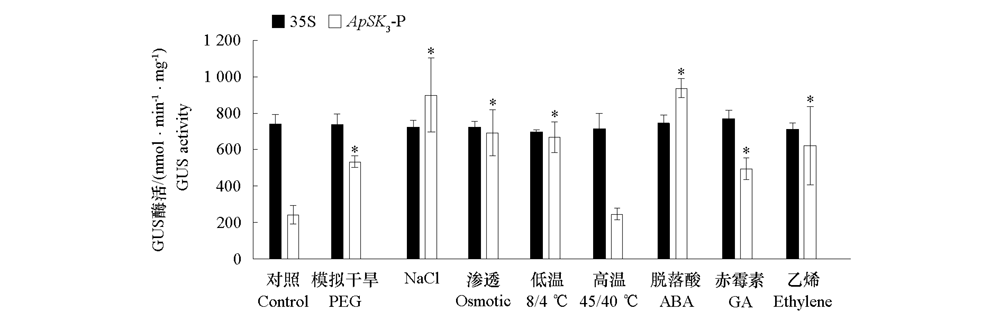
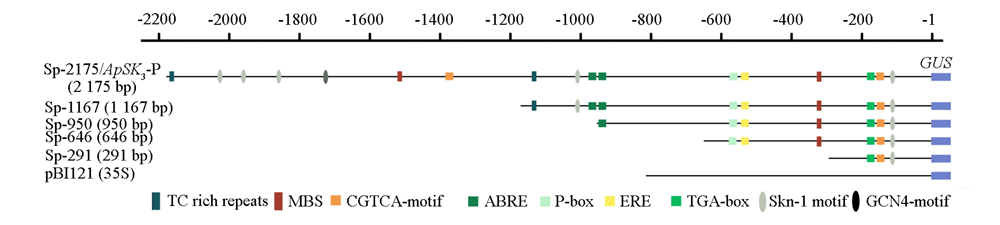
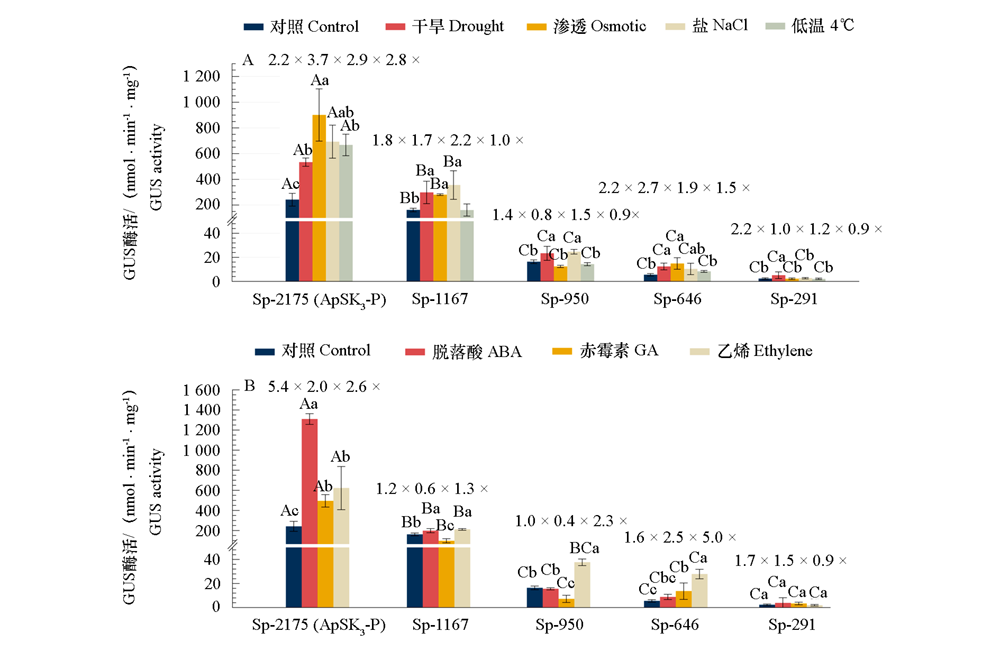
| [1] |
Abe H, Urao T, Ito T, Seki M, Shinozaki K, Yamaguchi-Shinozaki K. 2003. Arabidopsis AtMYC2(bHLH)and AtMYB2(MYB)function as transcriptional activators in abscisic acid signaling. The Plant Cell, 15 (1):63-78.
doi: 10.1105/tpc.006130
URL
|
| [2] |
Abedini R, GhaneGolmohammadi F, PishkamRad R, Pourabed E, Jafarnezhad A, Shobbar Z S, Shahbazi M. 2017. Plant dehydrins:shedding light on structure and expression patterns of dehydrin gene family in barley. Journal of Plant Research, 130 (4):747-763.
doi: 10.1007/s10265-017-0941-5
pmid: 28389925
|
| [3] |
Allagulova C R, Gimalov F R, Shakirova F M, Vakhitov V A. 2003. The plant dehydrins:structure and putative functions. Biochemistry(Moscow), 68 (9):945-51.
|
| [4] |
Bradford M M. 1976. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry, 72 (s 1-2):248-254.
doi: 10.1016/0003-2697(76)90527-3
URL
|
| [5] |
Chen R G, Jing H, Guo W L, Wang S B, Ma F, Pan B G, Gong Z H. 2015. Silencing of dehydrin CaDHN1 diminishes tolerance to multiple abiotic stresses in Capsicum annuum L. Plant Cell Reports, 34 (12):2189-2200.
doi: 10.1007/s00299-015-1862-1
URL
|
| [6] |
Guo X Y, Liu D F, Chong K. 2018. Cold signaling in plants:insights into mechanisms and regulation. Journal of Integrative Plant Biology, 60 (9):7-18.
|
| [7] |
Hernandez-Garcia C M, Finer J J. 2014. Identification and validation of promoters and cis-acting regulatory elements. Plant Science, 217-218 (1):109-119.
doi: 10.1016/j.plantsci.2013.12.007
URL
|
| [8] |
Hobo T, Asada M, Kowyama Y, Hattori T. 1999. ACGT‐containing abscisic acid response element(ABRE)and coupling element 3(CE3)are functionally equivalent. The Plant Journal, 19 (6):679-689.
doi: 10.1046/j.1365-313x.1999.00565.x
URL
|
| [9] |
Hundertmark M, Hincha D K. 2008. LEA(Late Embryogenesis Abundant)proteins and their encoding genes in Arabidopsis thaliana. BMC Genomics, 9 (1):118.
doi: 10.1186/1471-2164-9-118
URL
|
| [10] |
Hwang S H, Lee I A, Yie S W, Hwang D J. 2008. Identification of an OsPR10a promoter region responsive to salicylic acid. Planta, 227 (5):1141-1150.
doi: 10.1007/s00425-007-0687-8
URL
|
| [11] |
Jia F J, Qi S D, Li H, Liu P, Li P C, Wu C G, Zheng C C, Huang J G. 2014. Overexpression of Late Embryogenesis Abundant 14enhances Arabidopsis salt stress tolerance. Biochemical and Biophysical Research Communications, 454 (4):505-511.
doi: 10.1016/j.bbrc.2014.10.136
URL
|
| [12] |
Kidokoro S, Watanabe K, Ohori T, Moriwaki T, Maruyama K, Mizoi J, Htwe N, Fujita Y, Sekita S, Shinozaki K, Yamaguchi-Shinozaki K. 2015. Soybean DREB1/CBF‐type transcription factors function in heat and drought as well as cold stress‐responsive gene expression. The Plant Journal, 81 (3):505-518.
doi: 10.1111/tpj.12746
pmid: 25495120
|
| [13] |
Kruger C, Berkowitz O, Stephan U W, Hell R. 2002. A metal-binding member of the late embryogenesis abundant protein family transports iron in the phloem of Ricinus communis L. Journal of Biological Chemistry, 277 (28):25062-25069.
doi: 10.1074/jbc.M201896200
URL
|
| [14] |
Liang D, Xia H, Wu S, Ma F W. 2012. Genome-wide identification and expression profiling of dehydrin gene family in Malus domestica. Molecular Biology Reports, 39 (12):10759-10768.
doi: 10.1007/s11033-012-1968-2
URL
pmid: 23053973
|
| [15] |
Liu H, Yu C Y, Li H X, Ouyang B, Wang T T, Zhang J H, Wang X, Ye Z B. 2015. Overexpression of ShDHN,a dehydrin gene from Solanum habrochaites enhances tolerance to multiple abiotic stresses in tomato. Plant Science, 231:198-211.
doi: 10.1016/j.plantsci.2014.12.006
URL
|
| [16] |
Liu Yang. 2014. Isolation and functional analysis of LEA protein genes, ZmDHN13,ZmLEA3 and ZmLEA5C in Zea mays[Ph. D. Dissertation]. Tai’an:Shandong Agricultural University. (in Chinese)
|
|
刘洋. 2014. 玉米LEA蛋白基因ZmDHN13、ZmLEA3和ZmLEA5C的分离与功能分析[博士论文]. 泰安: 山东农业大学.
|
| [17] |
Lü A M, Fan N N, Xie J P, Yuan S L, An Y, Zhou P. 2017. Expression of CdDHN4,a novel YSK2-type dehydrin gene from bermudagrass,responses to drought stress through the ABA-dependent signal pathway. Frontiers in Plant Science, 8:748.
doi: 10.3389/fpls.2017.00748
URL
|
| [18] |
Ma Jie, Liu Cui-fang, Li Ling-zhi, Xiang Jian-hua, Chen Xin-bo. 2012. Progress in research of dehydrin response to abiotic stress. Journal of Biology, 29 (1):71-74. (in Chinese)
|
|
马杰, 刘翠芳, 李灵之, 向建华, 陈信波. 2012. 非生物胁迫下植物脱水素的研究进展. 生物学杂志, 29 (1):71-74.
|
| [19] |
Nakashima K, Kiyosue T, Yamaguchi-Shinozaki K, Shinozaki K. 1997. A nuclear gene,erd1,encoding a chloroplast-targeted Clp protease regulatory subunit homolog is not only induced by water stress but also developmentally up-regulated during senescence in Arabidopsis thaliana. The Plant Journal, 12 (4):851-861.
doi: 10.1046/j.1365-313X.1997.12040851.x
URL
|
| [20] |
Pellegrineschi A, Reynolds M, Pacheco M, Brito R M, Almeraya R, Yamaguchi-Shinozaki K, Hoisington D. 2004. Stress-induced expression in wheat of the Arabidopsis thaliana DREB1A gene delays water stress symptoms under greenhouse conditions. Genome, 47 (3):493-500.
pmid: 15190366
|
| [21] |
Qian Gang. 2007. Genotypic variability in sequences and expression of LEA2/LEA3 genes in Tibetan Hulless Barley,Hordeum vulgare ssp. vulgare,associated with resistance to water deficit[Ph. D. Dissertation]. Chengdu:Chengdu Institute of Biology,the Chinese Academy of Sciences. (in Chinese)
|
|
钱刚. 2007. 不同抗旱性青稞LEA2/LEA3蛋白基因的克隆与表达[博士论文]. 成都: 中国科学院研究生院.
|
| [22] |
Robertson M, Cuming A C, Chandler P M. 1995. Sequence analysis and hormonal regulation of a dehydrin promoter from barley, Hordeum vulgare. Physiologia Plantarum, 94 (3):470-478.
doi: 10.1111/ppl.1995.94.issue-3
URL
|
| [23] |
Rorat T. 2007. Plant dehydrins——Tissue location,structure and function. Cellular & Molecular Biology Letters, 11 (4):536-556.
|
| [24] |
Rorat T, Grygorowicz W J, Irzykowski W, Rey P. 2004. Expression of KS-type dehydrins is primarily regulated by factors related to organ type and leaf developmental stage during vegetative growth. Planta, 218 (5):878-885.
URL
pmid: 14685858
|
| [25] |
Saibi W, Feki K, Ben Mahmoud R, Brini F. 2015. Durum wheat dehydrin(DHN-5)confers salinity tolerance to transgenic Arabidopsis plants through the regulation of proline metabolism and ROS scavenging system. Planta, 242 (5):1187-1194.
doi: 10.1007/s00425-015-2351-z
URL
|
| [26] |
Swarup R, Parry G, Graham N, Allen T, Bennett M. 2002. Auxin cross-talk:integration of signalling pathways to control plant development. Plant Molecular Biology, 49 (3-4):411-426.
|
| [27] |
Uno Y, Furihata T, Abe H, Yoshida R, Shinozaki K, Yamaguchi-Shinozaki K. 2000. Arabidopsis basic leucine zipper transcription factors involved in an abscisic acid-dependent signal transduction pathway under drought and high-salinity conditions. Proceedings of the National Academy of Sciences of the United States of America, 97 (21):11632-11637.
|
| [28] |
Vasil V, Marcotte W R,Jr, Rosenkrans L, Cocciolone S M, Vasil I K, Quatrano R S, McCarty D R. 1995. Overlap of Viviparous1( VP1)and abscisic acid response elements in the Em promoter:G-box elements are sufficient but not necessary for VP1 transactivation. Plant Cell, 7:1511-1518.
pmid: 8589631
|
| [29] |
Wang C H, Gao G, Cao S X, Xie Q J, Qi H Y. 2019. Isolation and functional validation of the CmLOX08 promoter associated with signalling molecule and abiotic stress responses in oriental melon,Cucumis melo var. makuwa Makino. BMC Plant Biology, 19 (1):75.
doi: 10.1186/s12870-019-1678-1
URL
|
| [30] |
Wang H T, Zhang Y M, Xiao N, Zhang G, Wang F, Chen X Y, Fang R X. 2020a. Rice GERMIN-LIKE PROTEIN 2-1 functions in seed dormancy under the control of abscisic acid and gibberellic acid signaling pathways. Plant Physiology, 183 (3):1157-1170.
doi: 10.1104/pp.20.00253
URL
|
| [31] |
Wang Y D, Chen G J, Lei J J, Cao B H, Chen C M. 2020b. Identification and characterization of a LEA-like Gene,CaMF5,specifically expressed in the anthers of male-fertile Capsicum annuum. Horticultural Plant Journal, 26 (1):39-48.
|
| [32] |
Wu Qiong. 2019. Study of ABA-ethylene interaction on the regulation of cherry tomato fruit ripening[Ph. D. Dissertation]. Hangzhou:Zhejiang University. (in Chinese)
|
|
吴琼. 2019. ABA-乙烯互作调控樱桃番茄果实成熟的效应与机理研究[博士论文]. 杭州:浙江大学.
|
| [33] |
Yamaguchi-Shinozaki K, Shinozaki K. 1994. A novel cis-acting element in an Arabidopsis gene is involved in responsiveness to drought,low-temperature,or high-salt stress. Plant Cell, 6 (2):251-264.
pmid: 8148648
|
| [34] |
Yamaguchi-Shinozaki K, Shinozaki K. 2005. Organization of cis-acting regulatory elements in osmotic- and cold-stress-responsive promoters. Trends in Plant Science, 10 (2):88-94.
pmid: 15708346
|
| [35] |
Yang W B, Zhang L S, Lv H, Li H, Zhang Y N, Xu Y, Yu J N. 2015. The K-segments of wheat dehydrin WZY 2 are essential for its protective functions under temperature stress. Frontiers Plant Science, 6:406.
|
| [36] |
Yang Z, Sheng J Y, Lv K, Ren L, Zhang D. 2019. Y2SK2 and SK3 type dehydrins from Agapanthus praecox can improve plant stress tolerance and act as multifunctional protectants. Plant Science, 284:143-160.
doi: 10.1016/j.plantsci.2019.03.012
URL
|
| [37] |
Zhang H F, Liu S Y, Ma J H, Wang X K, Haq S U, Meng Y C, Zhang Y M, Chen R G. 2020. CaDHN4,a salt and cold stress-responsive dehydrin gene from pepper decreases abscisic acid sensitivity in Arabidopsis. International Journal of Molecular Sciences, 21 (1):26.
doi: 10.3390/ijms21010026
URL
|
| [38] |
Zhao Y, Wang Y, Liu Q, Zhai Y, Zhao Y, Zhang M J. 2017. Cloning of a new LEA 1 gene promoter from soybean and functional analysis in transgenic tobacco. Plant Cell Tissue and Organ Culture, 130 (4-5):1-13.
doi: 10.1007/s11240-017-1199-2
URL
|
| [39] |
Zhu K J, Wu Q J, Huang Y, Ye J L, Xu Q, Deng X X. 2020. Genome-wide characterization of cis-acting elements in the promoters of key carotenoid pathway genes from the main species of genus Citrus. Horticultural Plant Journal, 6 (6):385-395.
doi: 10.1016/j.hpj.2020.10.003
URL
|
| [40] |
Zhu W N, Zhang D P, Lu X X, Zhang L S, Yu Z Y, Lv H, Zhang H M. 2014. Characterisation of an SKn-type dehydrin promoter from wheat and its responsiveness to various abiotic and biotic stresses. Plant Molecular Biology Reporter, 32 (3):664-678.
doi: 10.1007/s11105-013-0681-1
URL
|