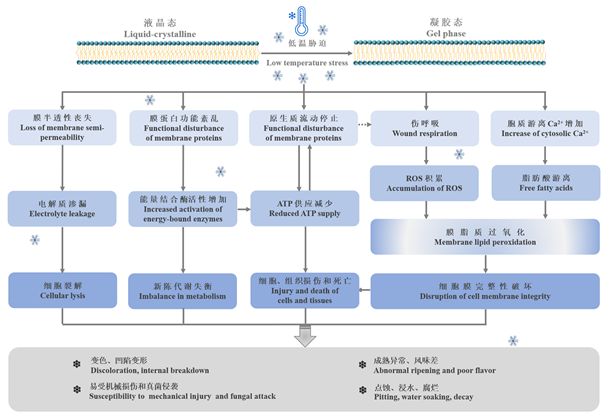
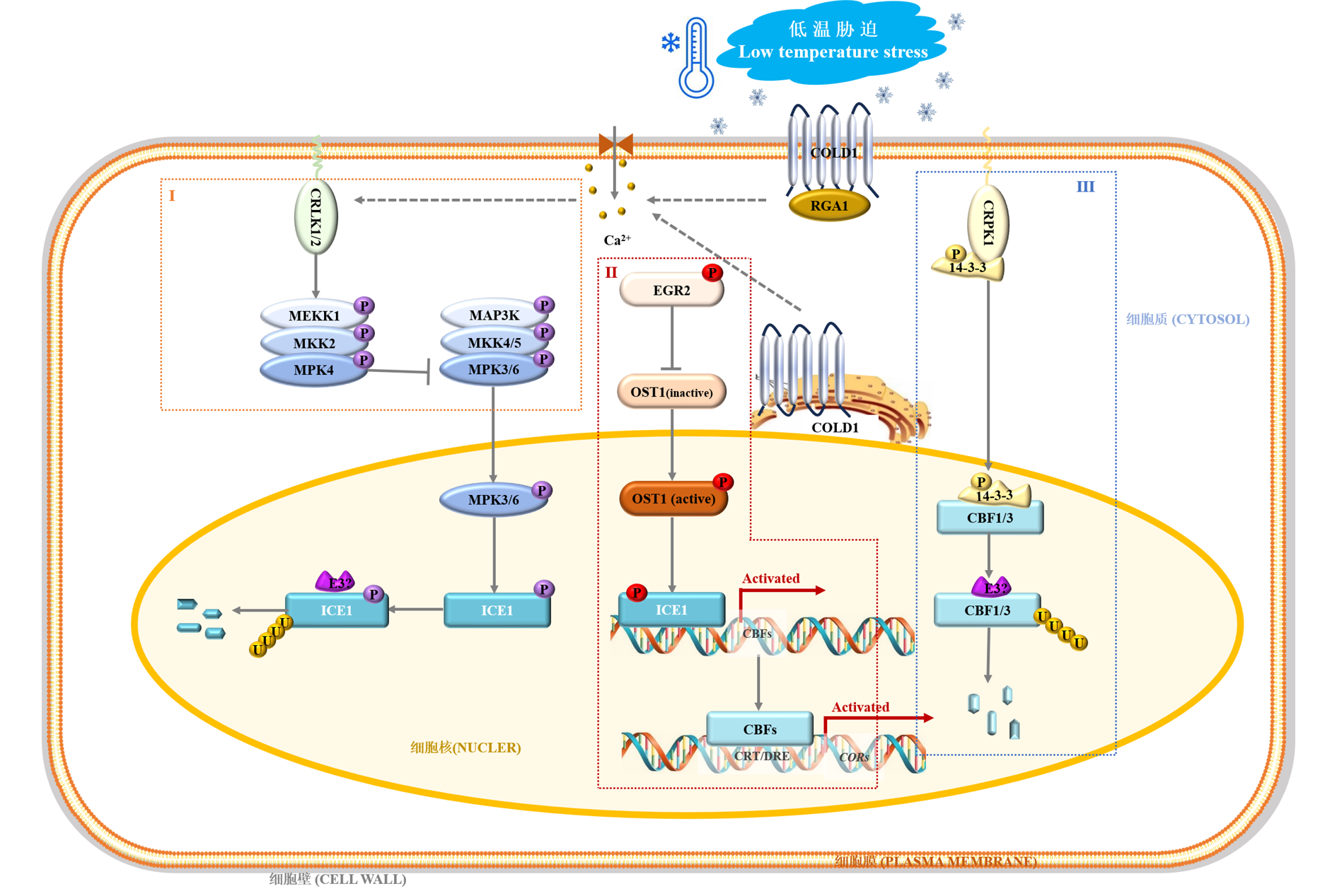
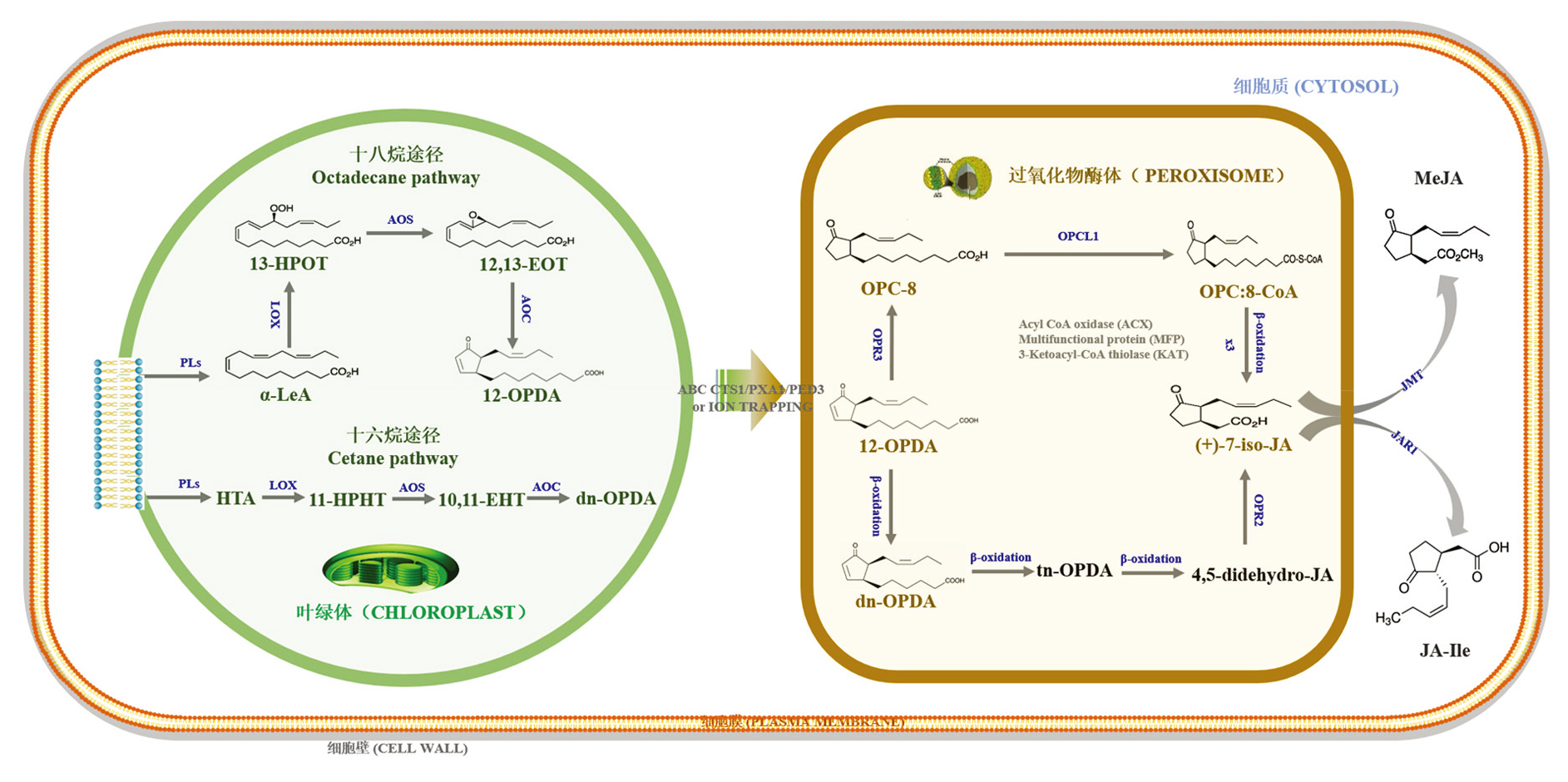
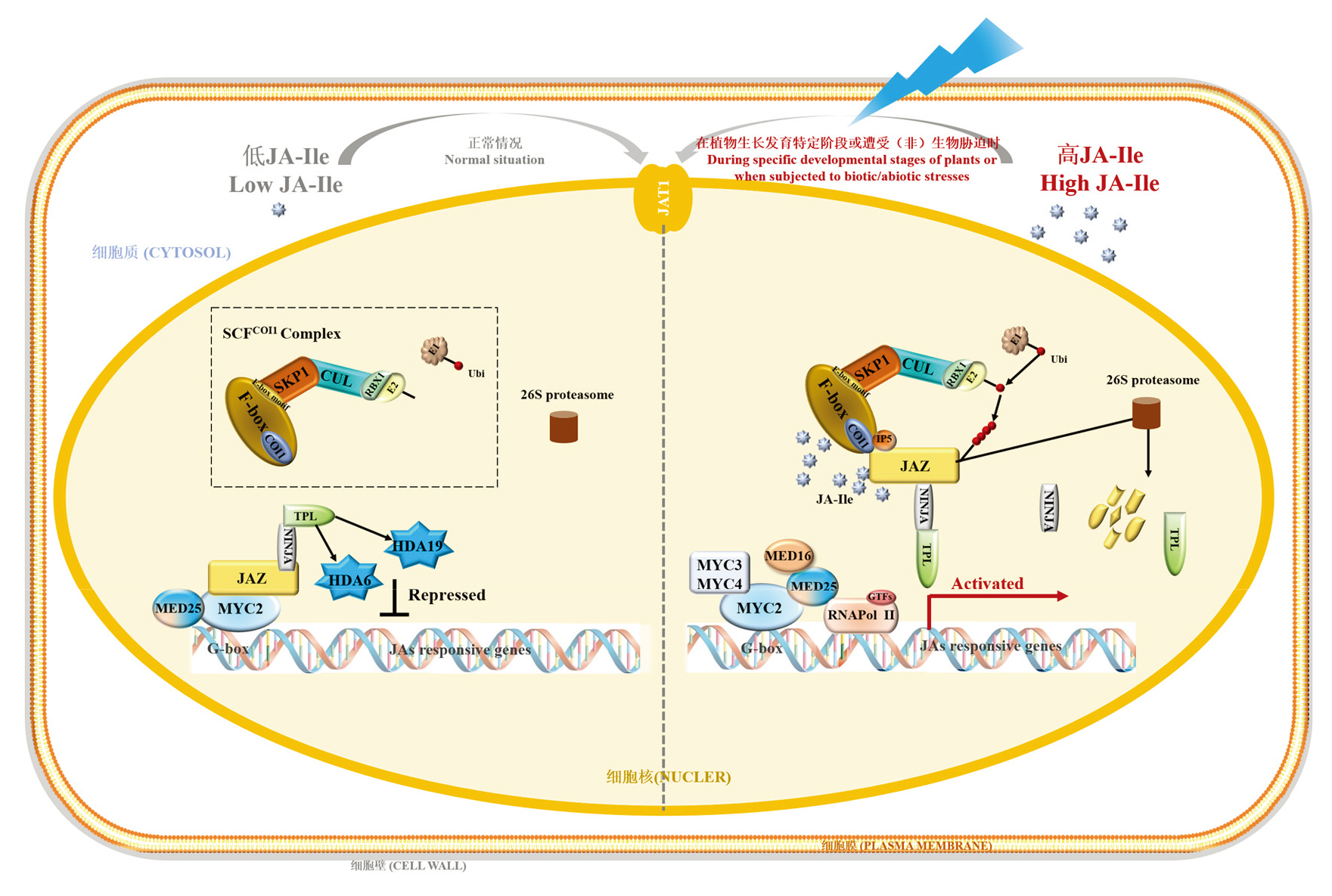
| [1] |
Ahmad M, Li J Z, Yang Q S, Jamil W, Teng Y W, Bai S L. 2019. Phylogenetic,molecular,and functional characterization of PpyCBF proteins in Asian pears(Pyrus pyrifolia). International Journal of Molecular Sciences, 20 (9):2074.
|
| [2] |
Ahmad P, Rasool S, Gul A, Sheikh S A, Akram N A, Ashraf M, Kazi A M, Gucel S. 2016. Jasmonates:Multifunctional roles in stress tolerance. Frontiers in Plant Science, 7:813.
|
| [3] |
Ai D, Zhao L, You C X, Han Y P, An J P. 2024. Apple SINA11-JAZ 2 module is involved in jasmonate signaling response. Journal of Integrative Plant Biology, 66 (7):1270-1273.
|
| [4] |
Ali M S, Baek K H. 2020. Jasmonic acid signaling pathway in response to abiotic stresses in plants. International Journal of Molecular Sciences, 21 (2):621.
|
| [5] |
An J P, Wang X F, Zhang X W, You C X, Hao Y J. 2021. Apple B-box protein BBX37 regulates jasmonic acid mediated cold tolerance through the JAZ-BBX37-ICE1-CBF pathway and undergoes MIEL1-mediated ubiquitination and degradation. New Phytologist, 229 (5):2707-2729.
|
| [6] |
An J P, Xu R R, Liu X, Su L, Yang K, Wang X F, Wang G L, You C X. 2022. Abscisic acid insensitive 4 interacts with ICE1 and JAZ proteins to regulate ABA signaling-mediated cold tolerance in apple. Journal of Experimental Botany, 73 (3):980-997.
|
| [7] |
Anjum N A, Ahmad I, Mohmood I, Pacheco M, Duarte A C, Pereira E, Umar Shahid, Ahmad A, Khan N A, Iqbal M, Prasad M N V. 2012. Modulation of glutatathione and its related enzymes in plants responses to toxic metals and metalloids-a review. Environmental and Experimental Botany, 75:307-324.
|
| [125] |
Roychoudhury A, Paul S, Basu S. 2013. Cross-talk between abscisic acid-dependent and abscisic acid-independent pathways during abiotic stress. Plant Cell Reports, 32 (7):985-1006.
doi: 10.1007/s00299-013-1414-5
pmid: 23508256
|
| [126] |
Roychowdhury R, Hada A, Biswas S, Mishra S, Prusty M R, Das S P, Ray S, Kumar A, Sarker U. 2024. Jasmonic acid(JA)in plant immune response:Unravelling complex molecular mechanisms and networking of defence signaling against pathogens. Journal of Plant Growth Regulation, 44:89-114.
|
| [127] |
Ruan J J, Zhou Y X, Zhou M L, Yan J, Khurshid M, Weng W F, Cheng J P, Zhang K X. 2019. Jasmonic acid signaling pathway in plants. International Journal of Molecular Sciences, 20 (10):2479.
|
| [128] |
Rubio S, Noriega X, Pérez F J. 2019. Abscisic acid(ABA)and low temperatures synergistically increase the expression of CBF/DREB 1 transcription factors and cold-hardiness in grapevine dormant buds. Annals of Botany, 123 (4):681-689.
|
| [129] |
Rutz J K, Borges C D, Zambiazi R C, da Rosa C G, da Silva M M. 2016. Elaboration of microparticles of carotenoids from natural and synthetic sources for applications in food. Food Chemistry, 202:324-333.
doi: 10.1016/j.foodchem.2016.01.140
pmid: 26920301
|
| [130] |
Sevillano L, Sanchez-Ballesta M T, Romojaro F, Flores F B. 2009. Physiological,hormonal and molecular mechanisms regulating chilling injury in horticultural species. Postharvest technologies applied to reduce its impact. Journal of the Science of Food and Agriculture, 89 (4):555-573.
|
| [131] |
Shah H M S, Singh Z, Hasan M U, Woodward A, Afrifa-Yamoah E. 2024. Preharvest methyl jasmonate application delays cell wall degradation and upregulates phenolic metabolism and antioxidant activities in cold stored raspberries. Food Chemistry, 462:141020.
|
| [8] |
Appelhagen I, Thiedig K, Nordholt N, Schmidt N, Huep G, Sagasser M, Weisshaar B. 2014. Update on transparent testa mutants from Arabidopsis thaliana:characterisation of new alleles from an isogenic collection. Planta, 240 (5):955-970.
doi: 10.1007/s00425-014-2088-0
pmid: 24903359
|
| [9] |
Ba L J, Kuang J F, Chen J Y, Lu W J. 2016. MaJAZ 1 attenuates the MaLBD5-mediated transcriptional activation of jasmonate biosynthesis gene MaAOC2 in regulating cold tolerance of banana fruit. Journal of Agricultural and Food Chemistry, 64:738-745.
|
| [10] |
Barrero-Gil J, Salinas J. 2018. Gene regulatory networks mediating cold acclimation:The CBF pathway. Advances in Experimental Medicine and Biology, 1081:3-22.
doi: 10.1007/978-981-13-1244-1_1
pmid: 30288701
|
| [11] |
Baswal A K, Dhaliwal H S, Gill K S, Ozturk B. 2023. Impact of modified atmospheric packaging films on health-promoting compounds in cold-stored‘Kinnow’mandarin(Citrus nobilis Lour × C. deliciosa Tenora)fruit. Erwerbs-Obstbau, 65:899-904.
|
| [12] |
Bolt S, Zuther E, Zintl S, Hincha D K, Schmülling T S. 2017. ERF 105 is a transcription factor gene of Arabidopsis thaliana required for freezing tolerance and cold acclimation. Plant,Cell & Environment, 40 (1):108-120.
|
| [13] |
Bonora M, Patergnani S, Rimessi A, De Marchi E, Suski J M, Bononi A, Giorgi C, Marchi S, Missiroli S, Poletti F, Wieckowski M R, Pinton P. 2012. ATP synthesis and storage. Purinergic Signal, 8 (3):343-357.
|
| [14] |
Cai H F, Han S, Yu M L, Ma R J, Yu Z F. 2020. The alleviation of methyl jasmonate on loss of aroma lactones correlated with ethylene biosynthesis in peaches. Journal of Food Science, 85 (8):2389-2397.
|
| [132] |
Shan W, Kuang J F, Lu W J, Chen J Y. 2014. Banana fruit NAC transcription factor MaNAC 1 is a direct target of MaICE1 and involved in cold stress through interacting with MaCBF1. Plant,Cell & Environment, 37 (9):2116-2127.
|
| [133] |
Sharma M, Laxmi A. 2016. Jasmonates:emerging players in controlling temperature stress tolerance. Frontiers in Plant Science, 6:11-29.
|
| [134] |
Shi H T, Chan Z L. 2014. AtHAP5A modulates freezing stress resistance in Arabidopsis independent of the CBF pathway. Plant Signaling & Behavior, 9 (7):e29109.
|
| [135] |
Shi Y T, Ding Y L, Yang S H. 2018. Molecular regulation of CBF signaling in cold acclimation. Trends in Plant Science, 23 (7):623-637.
doi: S1360-1385(18)30086-4
pmid: 29735429
|
| [136] |
Song Y, Liu L J, Feng Y H, Wei Y Z, Yue X L, He W L, Zhang H, An L Z. 2015. Chilling-and freezing-induced alterations in cytosine methylation and its association with the cold tolerance of an alpine subnival plant,Chorispora bungeana. PLoS ONE, 10 (8):e0135485.
|
| [137] |
Song Z Y, Lai X Y, Chen H C, Wang L H, Pang X Q, Hao Y W, Lu W J, Chen W X, Zhu X Y, Li X P. 2022. Role of MaABI5-like in abscisic acid-induced cold tolerance of‘Fenjiao’banana fruit. Horticulture Research, 9:uhac130.
|
| [138] |
Sood M. 2023. Jasmonates:“The master switch”for regulation of developmental and stress responses in plants. Journal of Plant Growth Regulation, 42:5247-5265.
|
| [139] |
Sun T, Rao S, Zhou X, Li L. 2022. Plant carotenoids:recent advances and future perspectives. Molecular Horticulture, 2 (1):3.
|
| [15] |
Cai Y T, Cao S F, Yang Z F, Zheng Y H. 2011. MeJA regulates enzymes involved in ascorbic acid and glutathione metabolism and improves chilling tolerance in loquat fruit. Postharvest Biology and Technology, 59 (3):324-326.
|
| [16] |
Cao K F, Wei Y Y, Chen Y, Jiang S, Chen X Y, Wang X X, Shao X F. 2021. PpCBF6 is a low-temperature-sensitive transcription factor that binds the PpVIN2 promoter in peach fruit and regulates sucrose metabolism and chilling injury. Postharvest Biology and Technology, 181:111681.
|
| [17] |
Cao S F, Cai Y T, Yang Z F, Zheng Y H. 2012. MeJA induces chilling tolerance in loquat fruit by regulating proline and γ-aminobutyric acid contents. Food Chemistry, 133 (4):1466-1470.
|
| [18] |
Cao S F, Zheng Y H, Wang K T, Jin P, Rui H J. 2009. Methyl jasmonate reduces chilling injury and enhances antioxidant enzyme activity in postharvest loquat fruit. Food Chemistry, 115 (4):1458-1463.
|
| [19] |
Cao S Q, Ye M, Jiang S T. 2005. Involvement of GIGANTEA gene in the regulation of the cold stress response in Arabidopsis. Plant Cell Reports, 24 (11):683-690.
|
| [20] |
Chan Z L, Wang Y P, Cao M J, Gong Y H, Mu Z X, Wang H Q, Hu Y L, Deng X, He X J, Zhu J K. 2016. RDM4 modulates cold stress resistance in Arabidopsis partially through the CBF-mediated pathway. New Phytologist, 209 (4):1527-1539.
|
| [21] |
Chen J J, Jin P, Li H H, Cai Y T, Zhao Y Y, Zheng Y H. 2012. Effects of low temperature storage on chilling injury and energy status in peach fruit. Transactions of the Chinese Society of Agricultural Engineering, 28:275-281.
|
| [140] |
Sun X M, Zhang L L, Wong D C J, Wang Y, Zhu Z F, Xu G Z, Wang Q F, Li S H, Liang Z C, Xin H P. 2019. The ethylene response factor VaERF 092 from amur grape regulates the transcription factor VaWRKY33,improving cold tolerance. The Plant Journal, 99 (5):988-1002.
|
| [141] |
Tang K, Zhao L, Ren Y Y, Yang S, Zhu J K, Zhao C Z. 2020. The transcription factor ICE 1 functions in cold stress response by binding to the promoters of CBF and COR genes. Journal of Integrative Plant Biology, 62 (3):258-263.
|
| [142] |
Tao X Y, Wu Q, Li J Y, Huang S Q, Cai Y, Mao L C, Luo Z S, Li L, Ying T J. 2022. Exogenous methyl jasmonate regulates phenolic compounds biosynthesis during postharvest tomato ripening. Postharvest Biology and Technology, 184:111760.
|
| [143] |
Theocharis A, Clement C, Barka E A. 2012. Physiological and molecular changes in plants grown at low temperatures. Planta, 235:1091-1105.
doi: 10.1007/s00425-012-1641-y
pmid: 22526498
|
| [144] |
Thines B, Katsir L, Melotto M, Niu Y J, Mandaokar A, Liu G H, Nomura K, He S Y, Howe G A, Browse J. 2007. JAZ repressor proteins are targets of the SCFCOI 1 complex during jasmonate signaling. Nature, 448 (7154):661-665.
|
| [145] |
Tian J X, Xie S Y, Zhang P, Wang Q, Li J K, Xu X B. 2022. Attenuation of postharvest peel browning and chilling injury of banana fruit by astragalus polysaccharides. Postharvest Biology and Technology, 184:111783.
|
| [22] |
Chen J, Kuang J F, Shan W, Wang J N, Xiao Y Y, Chen J Y, Lu W J. 2014. Molecular characterization of a cold-responsive RING-H 2 finger gene from banana fruit and its interaction with MaMYC2a. Postharvest Biology and Technology, 98:48-55.
|
| [23] |
Chen J, Li Y X, Li F F, Wu Q. 2018. Banana MaABI5 is involved in ABA-induced cold tolerance through interaction with a RING E3 ubiquitin ligase, MaC3HC4-1. Scientia Horticulturae, 237:239-246.
|
| [24] |
Chen Ming, Zhang Jieru, Yang Hangyun, Wang Yinbao, Zheng Zhiyuan, Zeng Jiaoke, Chen Jinyin, Fu Yongqi, Xiang Miaolian. 2024. Induction mechanism of methyl jasmonate on postharvest navel orange fruit against blue mold. Acta Horticulturae Sinica, 51 (9):2183-2194.
doi: 10.16420/j.issn.0513-353x.2023-0564
|
|
陈明, 张洁茹, 杨航云, 王印宝, 郑致远, 曾教科, 陈金印, 付永琦, 向妙莲. 2024. 茉莉酸甲酯诱导采后脐橙抗青霉病的机制研究. 园艺学报, 51 (9):2183-2194.
|
| [25] |
Chen Y M, Hoehenwarter W. 2019. Rapid and reproducible phosphopeptide enrichment by tandem metal oxide affinity chromatography:application to boron deficiency induced phosphoproteomics. The Plant Journal, 98 (2):370-384.
|
| [26] |
Chen Y M, Wang Y, Yang J, Zhou W B, Dai S J. 2021. Exploring the diversity of plant proteome. Journal of Integrative Plant Biology, 63 (7):1197-1210.
doi: 10.1111/jipb.13087
|
| [27] |
Chini A, Monte I, Zamarreno A M, Hamberg M, Lassueur S, Reymond P, Weiss S, Stintzi A, Schaller A, Porzel A, García-Mina J M, Solano R. 2018. An OPR3-independent pathway uses 4,5-didehydrojasmonate for jasmonate synthesis. Nature Chemical Biology, 14 (2):171-178.
doi: 10.1038/nchembio.2540
pmid: 29291349
|
| [28] |
Delgado C, Mora-Poblete F, Ahmar S, Chen J T, Figueroa C R. 2021. Jasmonates and plant salt stress:molecular players,physiological effects,and improving tolerance by using genome-associated tools. International Journal of Molecular Sciences, 22 (6):3082.
|
| [146] |
Vichaiya T, Faiyue B, Rotarayanont S, Uthaibutra J, Saengnil K. 2022. Exogenous trehalose alleviates chilling injury of‘Kim Ju’guava by modulating soluble sugar and energy metabolisms. Scientia Horticulturae, 301:111138.
|
| [147] |
Vithana M D K, Singh Z, Ul Hasan M. 2024. Pre-and post-harvest elicitation with methyl jasmonate and salicylic acid followed by cold storage synergistically improves red colour development and health-promoting compounds in blood oranges. Plant Growth Regulation, 43:1657-1671.
|
| [148] |
Vuolo M M, Lima V S, Junior M R M. 2019. Phenolic compounds:structure,classification,and antioxidant power//Maira Rubi Segura Campos. Bioactive Compounds. Woodhead Publishing:33-50.
|
| [149] |
Wan S, Xin X F. 2022. Regulation and integration of plant jasmonate signaling:a comparative view of monocot and dicot. Journal of Genetics and Genomics, 49 (8):704-714.
|
| [150] |
Wang C Y. 1989. Chilling injury of fruits and vegetables. Food Reviews International, 5:209-236.
|
| [151] |
Wang L, Chen S C, Shao J W, Zhang C, Mei L, Wang K, Jin P, Zheng Y H. 2022a. Hydrogen sulfide alleviates chilling injury in peach fruit by maintaining cell structure integrity via regulating endogenous H2S,antioxidant and cell wall metabolisms. Food Chemistry, 391:133283.
|
| [152] |
Wang L, Liu R, Yue Y T, Yu M, Zheng Y H, Zhang H. 2022b. Preservation treatment with methyl jasmonate alleviates chilling injury disorder in pear fruit by regulating antioxidant system and energy status. Journal of Food Processing and Preservation, 46 (1):e16152.
|
| [153] |
Wang L B, Baldwin E A, Bai J. 2016. Recent advance in aromatic volatile research in tomato fruit:the metabolisms and regulations. Food and Bioprocess Technology, 9:203-216.
|
| [154] |
Wang L B, Sun X X, Luo W Q, Qian C L. 2018. Roles of C-repeat binding factors-dependent signaling pathway in jasmonates-mediated improvement of chilling tolerance of postharvest horticultural commodities. Journal of Food Quality, Doi: 10.1155/2018/8517018.
|
| [29] |
Ding C K, Wang C Y, Gross K C, Smith D L. 2002. Jasmonate and salicylate induce the expression of pathogenesis-related-protein genes and increase resistance to chilling injury in tomato fruit. Planta, 214:895-901.
|
| [30] |
Ding F, Wang C, Xu N, Wang M L, Zhang S X. 2021. Jasmonic acid-regulated putrescine biosynthesis attenuates cold-induced oxidative stress in tomato plants. Scientia Horticulturae, 288:110373.
|
| [31] |
Ding F, Wang C, Zhang S X, Wang M L. 2022. A jasmonate-responsive glutathione S-transferase gene SlGSTU24 mitigates cold-induced oxidative stress in tomato plants. Scientia Horticulturae, 303:111231.
|
| [32] |
Ding F, Wang X Z, Li Z Y, Wang M L. 2023. Jasmonate positively regulates cold tolerance by promoting ABA biosynthesis in tomato. Plants, 12:60.
|
| [33] |
Ding Y L, Li H, Zhang X Y, Xie Q, Gong Z Z, Yang S H. 2015. OST 1 kinase modulates freezing tolerance by enhancing ICE1 stability in Arabidopsis. Developmental Cell, 32 (3):278-289.
|
| [34] |
Ding Y L, Lv J, Shi Y, Gao J, Hua J, Song C, Gong Z Z, Yang S H. 2019. EGR2 phosphatase regulates OST 1 kinase activity and freezing tolerance in Arabidopsis. EMBO Journal, 38 (1):e99819.
|
| [35] |
Divya P, Puthusseri B, Savanur M A, Lokesh V, Neelwarne B. 2018. Effects of methyl jasmonate and carotenogenic inhibitors on gene expression and carotenoid accumulation in coriander(Coriandrum sativum L.)foliage. Food Research International, 111:11-19.
|
| [36] |
Dong N Q, Lin H X. 2021. Contribution of phenylpropanoid metabolism to plant development and plant-environment interactions. Journal of Integrative Plant Biology, 63:180-209.
|
| [37] |
Duan W Y, Yang C, Cao X M, Zhang C, Liu H R, Chen K S, Li X, Zhang B. 2022. Transcriptome and DNA methylome analysis reveal new insights into methyl jasmonate-alleviated chilling injury of peach fruit after cold storage. Postharvest Biology and Technology, 189:111915.
|
| [38] |
Eremina M, Rozhon W, Poppenberger B. 2016. Hormonal control of cold stress responses in plants. Cellular And Molecular Life Sciences, 73 (4):797-810.
doi: 10.1007/s00018-015-2089-6
pmid: 26598281
|
| [39] |
Faizy A H, Ozturk B, Aglar E, Yıldız K. 2021. Role of methyl jasmonate application regime on fruit quality and bioactive compounds of sweet cherry at harvest and during cold storage. Journal of Food Processing and Preservation, 45 (10):e15882.
|
| [40] |
Fernández-Calvo P, Chini A, Fernández-Barbero G, Chico J M, Gimenez-Ibanez S, Geerinck J, Eeckhout D, Schweizer F, Godoy M, Franco-Zorrilla J M, Pauwels L, Witters E, Puga M I, Paz-Ares J, Goossens A, Reymond P, Jaeger G D, Solano R. 2011. The Arabidopsis bHLH transcription factors MYC3 and MYC 4 are targets of JAZ repressors and act additively with MYC2 in the activation of jasmonate responses. The Plant Cell, 23 (2):701-715.
doi: 10.1105/tpc.110.080788
pmid: 21335373
|
| [41] |
Fonseca S, Chico J M, Solano R. 2009. The jasmonate pathway:the ligand,the receptor and the core signaling module. Current Opinion in Plant Biology, 12 (5):539-547.
doi: 10.1016/j.pbi.2009.07.013
pmid: 19716757
|
| [42] |
Forman H J, Zhang H Q, Rinna A. 2009. Glutathione:overview of its protective roles,measurement,and biosynthesis. Molecular Aspects of Medicine, 30 (1-2):1-12.
doi: 10.1016/j.mam.2008.08.006
pmid: 18796312
|
| [43] |
Fung R W M, Wang C Y, Smith D L, Gross K C, Tao Y, Tian M. 2006. Characterization of alternative oxidase(AOX) gene expression in response to methyl salicylate and methyl jasmonate pre-treatment and low temperature in tomatoes. Journal of Plant Physiology, 163:1049-1060.
|
| [44] |
Gao H, Lu Z M, Yang Y, Wang D N, Yang T, Cao M M, Cao W. 2018. Melatonin treatment reduces chilling injury in peach fruit through its regulation of membrane fatty acid contents and phenolic metabolism. Food Chemistry, 245:659-666.
doi: S0308-8146(17)31637-0
pmid: 29287423
|
| [45] |
Gimenez-Ibanez S, Boter M, Solano R. 2015. Novel players fine-tune plant trade-offs. Essays in Biochemistry, 58:83-100.
doi: 10.1042/bse0580083
pmid: 26374889
|
| [46] |
González-Guzmán M, Apostoiova N, Belles J M, Barrero J M, Piqueras P, Ponce M R, Micol J L, Serrano R, Rodriguez P L. 2022. The short-chain alcohol dehydrogenase ABA 2 catalyzes the conversion of xanthoxin to abscisic aldehyde. The Plant Cell, 14 (8):1833-1846.
|
| [47] |
Goossens J, Fernández-Calvo P, Schweizer F, Goossens A. 2016. Jasmonates:signal transduction components and their roles in environmental stress responses. Plant Molecular Biology, 91 (6):673-689.
doi: 10.1007/s11103-016-0480-9
pmid: 27086135
|
| [48] |
Goswami A K, Maurya N K, Goswami S, Bardhan K, Singh S K, Prakash J, Pradhan S, Kumar A, Chinnusamy V, Kumar P, Sharma R M, Sharma S, Bisht D S, Kumar C. 2022. Physio-biochemical and molecular stress regulators and their crosstalk for low-temperature stress responses in fruit crops:a review. Frontiers in Plant Science, 13:1022167.
|
| [49] |
Griffiths G. 2020. Jasmonates:biosynthesis,perception and signal transduction. Essays in Biochemistry, 64 (3):501-512.
doi: 10.1042/EBC20190085
pmid: 32602544
|
| [50] |
Guo W L, Chen R G, Gong Z H, Yin Y X, Ahmedand S S, He Y M. 2012. Exogenous abscisic acid increases antioxidant enzymes and related gene expression in pepper(Capsicum annuum)leaves subjected to chilling stress. Genetics and Molecular Research, 11 (4):4063-4080.
doi: 10.4238/2012.September.10.5
pmid: 23079969
|
| [51] |
Guo X Y, Liu D F, Chong K. 2018. Cold signaling in plants:insights into mechanisms and regulation. Journal of Integrative Plant Biology, 60 (9):745-756.
|
| [52] |
Habibi F, Ramezanian A, Rahemi M, Eshghi S, Guillén F, Serrano M, Valero D. 2019. Postharvest treatments with γ-aminobutyric acid,methyl jasmonate,or methyl salicylate enhance chilling tolerance of blood orange fruit at prolonged cold storage. Journal of the Science of Food Agriculture, 99 (14):6408-6417.
|
| [53] |
Hardenburg R E, Watada A E, Yang C. 1990. The commercial storage of fruits,vegetables,and florist and nursery stocks// USDA agriculture handbook(No. 66). Washington: U.S. Government Printing Office:14-16.
|
| [54] |
Hasanuzzaman M, Bhuyan M H M B, Zulfiqar F, Raza A, Mohsin S M, Mahmud J A, Fujita M, Fotopoulos V. 2020. Reactive oxygen species and antioxidant defense in plants under abiotic stress:Revisiting the crucial role of a universal defense regulator. Antioxidants, 9:681.
|
| [55] |
Hasanuzzaman M, Fujita M. 2022. Plant oxidative stress:biology,physiology and mitigation. Plants, 11 (9):11-85.
|
| [56] |
Hemsley P A, Hurst C H, Kaliyadasa E, Lamb R, Knight M R, De Cothi E A, Steele J F, Knight H. 2014. The Arabidopsis mediator complex subunits MED16,MED14,and MED 2 regulate mediator and RNA polymerase II recruitment to CBF-responsive cold-regulated genes. The Plant Cell, 26 (1):465-484.
doi: 10.1105/tpc.113.117796
pmid: 24415770
|
| [57] |
Holthuis J C M, Menon A K. 2023. Lipid landscapes and pipelines in membrane homeostasis. Nature Reviews Molecular Cell Biology, 25 (3):194-213.
|
| [58] |
Howe G A, Major I T, Koo A J. 2018. Modularity in jasmonate signaling for multistress resilience. Annual Review of Plant Biology, 69:387-415.
doi: 10.1146/annurev-arplant-042817-040047
pmid: 29539269
|
| [59] |
Hu Y, Jiang Y J, Han X, Wang H P, Pan J J, Yu D Q. 2017. Jasmonate regulates leaf senescence and tolerance to cold stress:crosstalk with other phytohormones. Journal of Experimental Botany, 68 (6):1361-1369.
|
| [60] |
Hu Y F, Zhang H J, Gu B, Zhang J X. 2022. The transcription factor VaMYC 2 from Chinese wild Vitis amurensis enhances cold tolerance of grape (V. vinifera) by up-regulating VaCBF1 and VaP5CS. Plant Physiology and Biochemistry, 192:218-229.
|
| [61] |
Hu Y R, Jiang L Q, Wang F, Yu D Q. 2013. Jasmonate regulates the inducer of CBF expression-C-repeat binding factor/DRE binding factor 1 cascade and freezing tolerance in Arabidopsis. The Plant Cell, 25:2907-2924.
|
| [62] |
Huang J Y, Zhao X B, Bürger M, Chory J, Wang X C. 2023. The role of ethylene in plant temperature stress response. Trends in Plant Science, 28 (7):808-824.
doi: 10.1016/j.tplants.2023.03.001
pmid: 37055243
|
| [63] |
Huang T, Liu G S, Zhu L S, Liu J L, Xiang Y, Xu X B, Zhang Z K. 2024. Mitigation of chilling injury in mango fruit by methyl jasmonate is associated with regulation of antioxidant capacity and energy homeostasis. Postharvest Biology and Technology, 211:112801.
|
| [64] |
Huang X Y, Wang C K, Zhao Y W, Sun C H, Hu D G. 2021. Mechanisms and regulation of organic acid accumulation in plant vacuoles. Horticulture Research, 8:227.
|
| [155] |
Wang L H, Chen H, Chen G Y, Luo G B, Shen X Y, Ouyang B, Bie Z L. 2024a. Transcription factor SlWRKY 50 enhances cold tolerance in tomato by activating the jasmonic acid signaling. Plant Physiology, 194 (2):1075-1090.
|
| [156] |
Wang M, Fan X, Ding F. 2023. Jasmonate:a hormone of primary importance for temperature stress response in plants. Plants(Basel), 12 (24):4080.
|
| [157] |
Wang X, Liu J W, Wang M T, Liu L L, Liu X, Zhao C Z. 2024b. FERONIA controls ABA-mediated seed germination via the regulation of CARK 1 kinase activity. Cell Reports, 43 (11):114843.
|
| [158] |
Wang Y, Deng L, Meng J R, Niu L, Pan L, Lu Z H, Cui G C, Wang Z Q, Zeng W F. 2021. Transcriptomic and metabolic analyses reveal the mechanism of ethylene production in stony hard peach fruit during cold storage. International Journal of Molecular Science, 22 (21):11308
|
| [159] |
Wang Y C, Xu H F, Liu W J, Wang N, Qu C Z, Jiang S H, Fang H C, Zhang Z Y, Chen X S. 2019a. Methyl jasmonate enhances apple’cold tolerance through the JAZ-MYC 2 pathway. Plant Cell,Tissue and Organ Culture(PCTOC), 136 (1):75-84.
|
| [160] |
Wang Y X, Gao L P, Wang Q, Zuo J H. 2019b. Low temperature conditioning combined with methyl jasmonate can reduce chilling injury in bell pepper. Scientia Horticulturae, 243:434-439.
|
| [161] |
Watson S A, McStay G P. 2020. Functions of cytochrome c oxidase assembly factors. International Journal of Molecular Sciences, 21 (19):7254.
|
| [162] |
Wei M, Zhuang Y, Li H, Li P H, Huo H Q, Shu D, Huang W Z, Wang S H. 2020. The cloning and characterization of hypersensitive to salt stress mutant,affected in quinolinate synthase,highlights the involvement of NAD in stress-induced accumulation of ABA and proline. The Plant Journal, 102:85-98.
|
| [163] |
Winter G, Todd C D, Trovato M, Forlani G, Funck D. 2015. Physiological implications of arginine metabolism in plants. Frontiers in Plant Science, 6:534.
doi: 10.3389/fpls.2015.00534
pmid: 26284079
|
| [164] |
Wu F M, Sun C L, Zhu Z Y, Deng L, Yu F F, Xie Q, Li C Y. 2025. A multiprotein regulatory module,MED16-MBR1&2,controls MED25 homeostasis during jasmonate signaling. Nature Communications, 16 (1):772.
|
| [165] |
Wu J X, Nadeem M, Galagedara L, Thomas R, Cheema M. 2022a. Recent insights into cell responses to cold stress in plants:signaling,defence,and potential functions of phosphatidic acid. Environmental and Experimental Botany, 203:105068.
|
| [166] |
Wu J X, Tang R, Fan K. 2024a. Recent advances in postharvest technologies for reducing chilling injury symptoms of fruits and vegetables:a review. Food Chemistry, 21:101080.
|
| [167] |
Wu J, Kamanga B M, Zhang W Y, Xu Y H, Xu L. 2022b. Research progress of aldehyde oxidases in plants. Peer J, 10:e13119.
|
| [168] |
Wu P, Li B, Liu Y, Bian Z, Xiong J X, Wang Y X, Zhu B Z. 2024b. Multiple physiological and biochemical functions of ascorbic acid in plant growth,development,and abiotic stress response. International Journal Molecular Sciences, 25 (3):1832.
|
| [65] |
Hwarari D, Guan Y L, Ahmad B, Movahedi A, Min T, Hao Z D, Lu Y, Chen J H, Yang L M. 2022. ICE-CBF-COR signaling cascade and its regulation in plants responding to cold stress. International Journal of Molecular Sciences, 23 (3):1549.
|
| [66] |
Ishikawa T, Maruta T, Yoshimura K, Smirnoff N. 2018. Biosynthesis and regulation of ascorbic acid in plants//Gupta D,Palma J M,Corpas F J. Antioxidants and Antioxidant Enzymes in Higher Plants. Springer:Inc Cham:163-179.
|
| [67] |
Jia Y X, Ding Y L, Shi Y T, Zhang X Y, Gong Z Z, Yang S H. 2016. The CBFs triple mutants reveal the essential functions of CBFs in cold acclimation and allow the definition of CBF regulons in Arabidopsis. New Phytologist, 212 (2):345-353.
|
| [68] |
Jin P, Zhu H, Wang J, Chen J J, Wang X L, Zheng Y H. 2013. Effect of methyl jasmonate on energy metabolism in peach fruit during chilling stress. Journal of the Science of Food and Agriculture, 93:1827-1832.
doi: 10.1002/jsfa.5973
pmid: 23208649
|
| [69] |
Jin P, Zhu H, Wang L, Shan T M, Zheng Y H. 2014. Oxalic acid alleviates chilling injury in peach fruit by regulating energy metabolism and fatty acid contents. Food Chemistry, 161:87-93.
doi: 10.1016/j.foodchem.2014.03.103
pmid: 24837925
|
| [70] |
Johannes F, Porcher E, Teixeira F K, Saliba-Colombani V, Simon M, Agier N, Bulski A, Albuisson J, Heredia F, Audigier P, Bouchez D, Dillmann C, Guerche P, Hospital F, Colot V. 2009. Assessing the impact of transgenerational epigenetic variation on complex traits. PLoS Genetics, 5 (6):e1000530.
|
| [169] |
Wu Q, Wu J J, Sun H J, Zhang D, Yu D Y. 2011. Sequence and expression divergence of the AOC gene family in soybean:insights into functional diversity for stress responses. Biotechnology Letters, 33 (7):1351-1359.
|
| [170] |
Xiao J Q, Zhou Y J, Xie Y Y, Li T T, Su X G, He J X, Jiang Y M, Zhu H, Qu H X. 2024. ATP homeostasis and signaling in plants. Plant Communications, 5 (4):100834.
|
| [171] |
Xie G F, Liu N, Zhang Y, Tan S, Xu Q X, Luo Z S. 2024. Postharvest MeJA maintains the shelf quality of kiwifruit after cold storage by regulating antioxidant capacity and activating the disease resistance. Postharvest Biology and Technology, 211:112827.
|
| [172] |
Xin Z, Browse J. 1998. Eskimo 1 mutants of Arabidopsis are constitutively freezing-tolerant. Proceedings of the National Academy of Sciences of the United States of America, 95 (13):7799-7804.
|
| [173] |
Yan F, Zhang D, Wang X, Liu C, Zhang F. 2022. Reduction of postharvest diseases of loquat fruit by serine protease and possible mechanisms involved. Scientia Horticulturae, 304:111246.
|
| [174] |
Yang H Q, Wu F H, Cheng J Y. 2011. Reduced chilling injury in cucumber by nitric oxide and the antioxidant response. Food Chemistry, 127 (3):1237-1242.
doi: 10.1016/j.foodchem.2011.02.011
pmid: 25214120
|
| [175] |
Yang L, Sun Q G, Geng B H, Shi J, Zhu H F, Sun Y M, Yang Q Y, Yang B, Guo Z F. 2023. Jasmonate biosynthesis enzyme allene oxide cyclase 2 mediates cold tolerance and pathogen resistance. Plant Physiology, 193 (2):1621-1634.
doi: 10.1093/plphys/kiad362
pmid: 37392433
|
| [176] |
Ye Y J, Xiao Y Y, Han Y C, Shan W, Fan Z Q, Xu Q G, Kuang J F, Lu W J, Lakshmanan P, Chen J Y. 2016. Banana fruit VQ motif-containing protein5 represses cold-responsive transcription factor MaWRKY26 involved in the regulation of JA biosynthetic genes. Scientific Reports, 6 (1):23632.
|
| [177] |
Yin X C, Ji S J, Cheng S C, Zhou Q, Zhou X, Luo M L, Ma M J, Hu M S, Wei B D. 2021. Methyl jasmonate alleviates the reduced release of aroma-related esters in‘Nanguo’pears by regulating ethylene biosynthesis and signal transduction. International Journal of Food Science & Technology, 56 (2):814-824.
|
| [178] |
Yu W Q, Jia X X, Shen L, Sheng J P. 2023. Ethylene perception influences the chilling injury and SlCBF1 gene expression during cold storage of tomato fruit. Postharvest Biology and Technology, 204:112434.
|
| [179] |
Yuan H, Zhang J, Nageswaran D, Li L. 2015. Carotenoid metabolism and regulation in horticultural crops. Horticulture Research, 2:15036.
doi: 10.1038/hortres.2015.36
pmid: 26504578
|
| [180] |
Yue P T, Jiang Z H, Sun Q, Wei R R, Yin Y Z, Xie Z Z, Larkin R M, Ye J L, Chai L J, Deng X X. 2023. Jasmonate activates a CsMPK6-CsMYC 2 module that regulates the expression of β-citraurin biosynthetic genes and fruit coloration in orange(Citrus sinensis). The Plant Cell, 35 (4):1167-1185.
|
| [181] |
Zhang P, Shao X F, Wei Y Y, Xu F, Wang H F. 2020. At-harvest fruit maturity affects sucrose metabolism during cold storage and is related to chilling injury in peach. Journal of Food Science and Technology, 57 (6):2000-2009.
doi: 10.1007/s13197-019-04232-4
pmid: 32431326
|
| [182] |
Zhang X B, Liu C J. 2015. Multifaceted regulations of gateway enzyme phenylalanine ammonia-lyase in the biosynthesis of phenylpropanoids. Molecular Plant, 8:17-27.
doi: 10.1016/j.molp.2014.11.001
pmid: 25578269
|
| [183] |
Zhang X H, Sheng J P, Li F J, Meng D M, Shen L. 2012. Methyl jasmonate alters arginine catabolism and improves postharvest chilling tolerance in cherry tomato fruit. Postharvest Biology and Technology, 64:160-167.
|
| [184] |
Zhang Y X, Zhang Z P, Ai Y X, Zhang H Z, Chen Y, Ye Y Q, Sun L, Shen H L, Cheng Q. 2024. CaAOS as a hub gene based on physiological and transcriptomic analyses of cold-resistant and cold-sensitive pepper cultivars.. International Journal of Biological Macromolecules, 276:133961.
|
| [185] |
Zhang Y, Zeng L. 2020. Crosstalk between ubiquitination and other post-translational protein modifications in plant immunity. Plant Communications, 1 (4):100041.
|
| [186] |
Zhao C Z, Wang P C, Si T, Hsu C C, Wang L, Zayed O, Yu Z P, Zhu Y F, Dong J, Tao W A, Zhu J K. 2017. MAP kinase cascades regulate the cold response by modulating ICE 1 protein stability. Developmental Cell, 43 (5):618-629.
|
| [187] |
Zhao M L, Wang J N, Shan W, Fan J G, Kuang J F, Wu K Q, Li X P, Chen W X, He F Y, Chen J Y, Lu W J. 2013. Induction of jasmonate signaling regulators MaMYC2s and their physical interactions with MaICE 1 in methyl jasmonate-induced chilling tolerance in banana fruit. Plant,Cell & Environment, 36 (1):30-51.
|
| [188] |
Zhao Y Y, Song C C, Brummell D A, Qi S N, Lin Q, Duan Y Q. 2021. Jasmonic acid treatment alleviates chilling injury in peach fruit by promoting sugar and ethylene metabolism. Food Chemistry, 338:128005.
|
| [189] |
Zheng P, Cao L, Zhang C, Fang X, Wang L, Miao M, Tang X, Liu Y, Cao S. 2023. The transcription factor MYB 43 antagonizes with ICE1 to regulate freezing tolerance in Arabidopsis. New Phytologist, 238 (6):2440-2459.
|
| [190] |
Zheng X Z, Gong M, Zhang Q D, Tan H Q, Li L P, Tang Y W, Li Z G, Peng M C, Deng W. 2022. Metabolism and regulation of ascorbic acid in fruits. Plants(Basel), 11 (12):1602.
|
| [191] |
Zhou Y, Liu X C, Liang X Y, Li H M, Lai J H, Liao Y R, Liu K D. 2024. Biochemical and metabolomics analyses reveal the mechanisms underlying ascorbic acid and chitosan coating mediated energy homeostasis in postharvest papaya fruit. Food Chemistry, 439:138-168.
|
| [192] |
Zhu J, Wang W S, Yan D W, Hong L W, Li T T, Gao X, Yang Y H, Ren F, Lu Y T, Yuan T T. 2023. CK 2 promotes jasmonic acid signaling response by phosphorylating MYC2 in Arabidopsis. Nucleic Acids Research, 51:619-630.
|
| [193] |
Zhu J H, Shi H Z, Lee B H, Damsz B, Cheng S, Stirm V, Zhu J K, Hasegawa P M, Bressan R A. 2004. An Arabidopsis homeodomain transcription factor gene,HOS9,mediates cold tolerance through a CBF-independent pathway. Proceedings of the National Academy of Sciences of the United States of America, 101 (26):9873-9878.
|
| [71] |
Jomova K, Valko M. 2013. Health protective effects of carotenoids and their interactions with other biological antioxidants. European Journal of Medicinal Chemistry, 70:102-110.
doi: 10.1016/j.ejmech.2013.09.054
pmid: 24141200
|
| [72] |
Kazan K, Manners J M. 2008. Jasmonate signaling:toward an integrated view. Plant Physiology, 146 (4):1459-1468.
|
| [73] |
Kerchev P I, Pellny T K, Vivancos P D, Kiddle G, Hedden P, Driscoll S, Vanacker H, Verrier P, Hancock R D, Foyer C H. 2011. The transcription factor ABI 4 is required for the ascorbic acid-dependent regulation of growth and regulation of jasmonate-dependent defense signaling pathways in Arabidopsis. The Plant Cell, 23 (9):3319-3334.
doi: 10.1105/tpc.111.090100
pmid: 21926335
|
| [74] |
Khan M, Hu J B, Dahro B, Ming R H, Zhang Y, Wang Y, Alhag A, Li C L, Liu J H. 2021. ERF108 from Poncirus trifoliata(L.)Raf. functions in cold tolerance by modulating raffinose synthesis through transcriptional regulation of PtrRafS. The Plant Journal:for Cell and Moiecular Biology, 108 (3):705-724.
|
| [75] |
Kim Y S, Lee M, Lee J H, Lee H J, Park C M. 2015. The unified ICE-CBF pathway provides a transcriptional feedback control of freezing tolerance during cold acclimation in Arabidopsis. Plant Molecular Biology, 89 (1-2):187-201.
|
| [76] |
Kitinoja L, Kader A A. 1995. Small scale postharvest handling practices-a manual for horticultural crops. USA: University of California-Davis.
|
| [77] |
Kooijman E E, Chupin V, Kruijff B D, Burger K N J. 2003. Modulation of membrane curvature by phosphatidic acid and lysophosphatidic acid J. Traffic, 4 (3):162-174.
doi: 10.1034/j.1600-0854.2003.00086.x
pmid: 12656989
|
| [78] |
Lee T M, Lur H S, Lin Y H, Chu C. 1996. Physiological and biochemical changes related to methyl jasmonate‐induced chilling tolerance of rice (Oryza sativa L.) seedlings. Plant,Cell & Environment, 19 (1):65-74.
|
| [194] |
Zhu L S, Ting H, Liu J L, Xu X B, Zhang Z K. 2024. Transcriptome analysis reveals the potential mechanism of methyl jasmonate alleviated ripening disorder in mango fruit at low temperature. Food Chemistry, 463:141093.
|
| [79] |
Li C, Xu M X, Cai X, Han Z G, Si J P, Chen D H. 2022. Jasmonate signaling pathway modulates plant defense,growth,and their trade-offs. International Journal of Molecular Medicine, 23 (7):39-45.
|
| [80] |
Li H, Ding Y L, Shi Y T, Zhang X Y, Zhang S Q, Gong Z Z, Yang S H. 2017a. MPK3-and MPK6-mediated ICE 1 phosphorylation negatively regulates ICE1 stability and freezing tolerance in Arabidopsis. Developmental Cell, 43 (5):630-642.
|
| [81] |
Li H, Ye K Y, Shi Y T, Cheng J K, Zhang X Y, Yang S H. 2017b. BZR1 positively regulates freezing tolerance via CBF-dependent and CBF-independent pathways in Arabidopsis. Molecular Plant, 10 (4):545-559.
|
| [82] |
Li L, Dou N, Zhang H, Wu C X. 2021a. The versatile GABA in plants. Plant Signaling & Behavior, 16 (3):1862565.
|
| [83] |
Li M Y, Yu G H, Cao C L, Liu P. 2021b. Metabolism,signaling,and transport of jasmonates. Plant Communications, 2 (5):100231.
|
| [84] |
Li P Y, Zheng X L, Liu Y, Zhu Y Y. 2014. Pre-storage application of oxalic acid alleviates chilling injury in mango fruit by modulating proline metabolism and energy status under chilling stress. Food Chemistry, 142:72-78.
doi: 10.1016/j.foodchem.2013.06.132
pmid: 24001814
|
| [85] |
Li T, XU Y X, Zhang L C, Ji Y L, Tan D M, Yuan H, Wang A D. 2017c. The jasmonate-activated transcription factor MdMYC 2 regulates ethylene response factor and ethylene biosynthetic genes to promote ethylene biosynthesis during apple fruit ripening. The Plant Cell, 29 (6):1316-1334.
|
| [86] |
Li Y L, Zhao Y T, Zhang Z C, He H, Shi L, Zhu X, Cui K B. 2022. Near-freezing temperature storage improves shelf-life and suppresses chilling injury in postharvest apricot fruit(Prunus armeniaca L.)by regulating cell wall metabolism. Food Chemistry, 387:132921.
|
| [87] |
Li Z L, Min D D, Fu X D, Zhao X M, Wang J H, Zhang X H, Li F J, Li X A. 2021c. The roles of SlMYC2 in regulating ascorbate-glutathione cycle mediated by methyl jasmonate in post-harvest tomato fruits under cold stress. Scientia Horticulturae, 288:110406.
|
| [88] |
Libeisson Y, Shorrosh B, Beisson F, Andersson M X, Arondel V, Bates P D, Baud S, Bird D, DeBono A, Durrett T P, Franke R B, Graham I A, Katayama K, Kelly A A, Larson T, Markham J E, Miquel M, Molina I, Nishida I, Rowland O, Samuels L, Schmid K M, Wada H, Welti R, Xu C C, Zallot R, Ohlrogge. 2010. Acyl-lipid metabolism. Arabidopsis Book, 8:e0161.
|
| [89] |
Lin Y X, Lin H T, Chen Y H, Wang H, Ritenour M A, Lin Y F. 2019. Hydrogen peroxide-induced changes in activities of membrane lipids-degrading enzymes and contents of membrane lipids composition in relation to pulp breakdown of longan fruit during storage. Food Chemistry, 297:124955.
|
| [90] |
Lin Z, Li Y, Wang Y, Liu X, Ma L, Zhang Z, Mu C, Zhang Y, Peng L, Xie S, Song C P, Shi H, Zhu J K, Wang P. 2021. Initiation and amplification of SnRK 2 activation in abscisic acid signaling. Nature Communications, 12 (1):2456.
|
| [91] |
Liu B, Seong K, Pang S H, Song J Q, Gao H, Wang C L, Zhai J Q, Zhang Y, Gao S, Li X D, Qi T C, Song S S. 2021a. Functional specificity,diversity,and redundancy of Arabidopsis JAZ family repressors in jasmonate and COI1-regulated growth,development,and defense. New Phytologist, 231 (4):1525-1545.
|
| [92] |
Liu W H, Wang H Y, Chen Y P, Zhu S Q, Chen M, Lan X Z, Chen G P, Liao Z H. 2017a. Cold stress improves the production of artemisinin depending on the increase in endogenous jasmonate. Biotechnology and Applied Biochemistry, 64 (3):305-314.
|
| [93] |
Liu W X, Feng Y, Yu S H, Fan Z Q, Li X L, Li J Y, Yin H F. 2021b. The flavonoid biosynthesis network in plants. International Journal Molecular Sciences, 22 (23):12824.
|
| [94] |
Liu Z Y, Jia Y X, Ding Y L, Shi Y T, Li Z, Guo Y, Gong Z Z, Yang S H. 2017b. Plasma membrane CRPK1-mediated phosphorylation of 14-3-3 proteins induces their nuclear import to fine-tune CBF signaling during cold response. Molecular Cell, 66 (1):117-128
|
| [95] |
Lu Z G, Wang X W, Lin X Y, Mostafa S, Zou H L, Wang L, Jin B. 2024. Plant anthocyanins:classification,biosynthesis,regulation,bioactivity,and health benefits. Plant Physiology Biochemistry, 217:109268.
|
| [96] |
Luo W, Huan Q, Xu Y Y, Qian W F, Chong K, Zhang J. 2022. Integrated global analysis reveals a vitamin E-vitamin K 1 sub-network,downstream of COLD1,underlying rice chilling tolerance divergence. Cell Reports, 36 (3):109397.
|
| [97] |
Lurie S. 2021. Genomic and transcriptomic studies on chilling injury in peach and nectarine. Postharvest Biology and Technology, 174:111444.
|
| [98] |
Lyons J M. 1973. Chilling injury in plants. Annual Review of Plant Biology, 24:445-466.
|
| [99] |
Ma M J, Zhu Z Q, Cheng S C, Zhou Q, Zhou X, Kong X M, Hu M S, Yin X C, Wei B D, Ji S J. 2020. Methyl jasmonate alleviates chilling injury by regulating membrane lipid composition in green bell pepper. Scientia Horticulturae, 266:109308.
|
| [100] |
Meesters C, Kombrink E. 2014. Jasmonic acid//Audenaert D,Overvoorde P. Plant chemical biology. Hoboken: Wiley:160-180.
|
| [101] |
Meyer K, Leube M P, Grill E. 1994. A protein phosphatase 2C involved in ABA signal transduction in Arabidopsis thaliana. Science, 264:1452-1455.
doi: 10.1126/science.8197457
pmid: 8197457
|
| [102] |
Min D D, Li F J, Ali M, Zhang X H, Liu Y G. 2023. Application of methyl jasmonate to control chilling tolerance of postharvest fruit and vegetables:A meta-analysis and eliciting metabolism review. Critical Reviews in Food Science and Nutrition, 64 (33):1-14.
doi: 10.1080/10408398.2023.2249097
pmid: 37615648
|
| [103] |
Min D D, Zhou J X, Li J Z, Ai W, Li Z L, Zhang X H, Fu X D, Zhao X M, Li F J, Li X A, Guo Y Y. 2021. SlMYC2 targeted regulation of polyamines biosynthesis contributes to methyl jasmonate-induced chilling tolerance in tomato fruit. Postharvest Biology and Technology, 174:111443.
|
| [104] |
Ming R, Zhang Y, Wang Y, Khan M, Dahro B, Liu J. 2021. The JA-responsive MYC2-BADH-like transcriptional regulatory module in poncirus trifoliata contributes to cold tolerance by modulation of glycine betaine biosynthesis. New Phytologist, 229:2730-2750.
doi: 10.1111/nph.17063
pmid: 33131086
|
| [105] |
Mirdehghan S H, Ghotbi F. 2014. Effects of salicylic acid,jasmonic acid,and calcium chloride on reducing chilling injury of pomegranate(Punica granatum L.) fruit. Journal of Agricultural Science and Technology, 16 (1):163-173.
|
| [106] |
Mittler R, Zandalinas S I, Fichman Y, Breusegem F V. 2022. Reactive oxygen species signaling in plant stress responses. Nature Reviews Molecular Cell Biology, 23:663-679.
|
| [107] |
Miura K, Shiba H, Ohta M, Kang S W, Sato A, Yuasa T, Iwaya Inoue M, Kamada H, Ezura H. 2012. SlICE1 encoding a MYC-type transcription factor controls cold tolerance in tomato,Solanum lycopersicum. Plant Biotechnology. 29 (3):253-260.
|
| [108] |
Mizoi J, Shinozaki K, Yamaguchi-Shinozaki K. 2012. AP2/ERF family transcription factors in plant abiotic stress responses. Biochimica et Biophysica Acta-Gene Regulatory Mechanisms, 1819 (2):86-96.
|
| [109] |
Muzaffar A, Chen Y S, Lee H T, Wu C C, Le T T, Liang J Z, Lu C H, Balasubramaniam H, Lo S F, Yu L C, Chan C H, Chen K T, Lee M H, Hsing Y I, Ho T H D, Yu S M. 2024. A newly evolved rice-specific gene JAUP1 regulates jasmonate biosynthesis and signaling to promote root development and multi-stress tolerance. Plant Biotechnology Journal, 22 (5):1417-1432.
doi: 10.1111/pbi.14276
pmid: 38193234
|
| [110] |
Nguyen T H, Goossens A, Lacchini E. 2022. Jasmonate:a hormone of primary importance for plant metabolism. Current Opinion in Plant Biology, 67:102-197.
|
| [111] |
Nurbekova Z, Srivastava S, Standing D, Kurmanbayeva A, Bekturova A, Soltabayeva A, Oshanova D, Turečková V, Strand M, Biswas M S, Mano J, Sagi M. 2021. Arabidopsis aldehyde oxidase 3,known to oxidize abscisic aldehyde to abscisic acid,protects leaves from aldehyde toxicity. The Plant Journal, 108 (5):1439-1455.
|
| [112] |
Olate E, Jimenez-Gomez J M, Holuigue L, Salinas J. 2018. NPR1 mediates a novel regulatory pathway in cold acclimation by interacting with HSFA1 factors. Nature Plants, 4 (10):811-823.
doi: 10.1038/s41477-018-0254-2
pmid: 30250280
|
| [113] |
Oliveira J A A, Salomão L C C, Siqueira D L D, Cecon P R. 2016. Cold tolerance of banana fruits of different cultivars. Revista Caatinga, 29 (3):629-641.
|
| [114] |
Park M H, Sangwanangkul P, Choi J W. 2018. Reduced chilling injury and delayed fruit ripening in tomatoes with modified atmosphere and humidity packaging. Scientia Horticulturae, 231:66-72.
|
| [115] |
Park S, Lee C M, Doherty C J, Gilmour S J, Kim Y S, Thomashow M F. 2015. Regulation of the Arabidopsis CBF regulon by a complex low-temperature regulatory network. Journal of Plant Pathology, 82 (2):193-207.
|
| [116] |
Patel B, Tandel Y N, Patel A H, Patel B L. 2016. Chilling injury in tropical and subtropical fruits:a cold storage problem and its remedies:a review. International Journal of Science,Environment and Technology, 5 (4):1882-1887.
|
| [117] |
Pattyn J, Vaughan-Hirsch J, Poel B V D. 2021. The regulation of ethylene biosynthesis:a complex multilevel control circuitry. New Phytologist, 229 (2):770-782.
|
| [118] |
Paull R E. 1990. Chilling injury of crops of tropical and subtropical origin//Wang J Y. Chilling injury of horticultural crops. Boca Raton: CRC Press:17-36.
|
| [119] |
Qi C D, Dong D H, Li Y F, Wang X W, Guo L Q, Liu L, Dong X N, Li X S, Yuan X S, Ren S X, Zhang N, Guo Y D. 2022. Heat shock-induced cold acclimation in cucumber through CsHSFA1d-activated JA biosynthesis and signaling. The Plant Journal, 111 (1):85-102.
|
| [120] |
Qi X N, Xiao Y Y, Fan Z Q, Chen J Y, Lu W J, Kuang J F. 2016. A banana fruit transcriptional repressor MaERF 10 interacts with MaJAZ3 to strengthen the repression of JA biosynthetic genes involved in MeJA-mediated cold tolerance. Postharvest Biology and Technology, 120:222-231.
|
| [121] |
Raza A, Charagh S, Zahid Z, Mubarik M S, Javed R, Siddiqui M H, Hasanuzzaman M. 2021. Jasmonic acid:a key frontier in conferring abiotic stress tolerance in plants. Plant Cell Reports, 40 (8):1513-1541.
doi: 10.1007/s00299-020-02614-z
pmid: 33034676
|
| [122] |
Raza A, Charagh S, Najafi-Kakavand S, Abbas S, Shoaib Y, Anwar S, Sharifi S, Lu G Y, Siddique K H M. 2023. Role of phytohormones in regulating cold stress tolerance:Physiological and molecular approaches for developing cold-smart crop plants. Plant Stress, 8:100152.
|
| [123] |
Repkina N, Ignatenko A, Holoptseva E, MiszalskI Z, Kaszycki P, Talanova V. 2021. Exogenous methyl jasmonate improves cold tolerance with parallel induction of two cold-regulated(COR)genes expression in Triticum aestivum L. Plants, 10 (7):1421.
|
| [124] |
Rodriguez P L, Benning G, Grill E. 1998. ABI2,a second protein phosphatase 2C involved in abscisic acid signal transduction in Arabidopsis. FEBS Letters, 421:185-190.
doi: 10.1016/s0014-5793(97)01558-5
pmid: 9468303
|